So the robust australopithecines have some interesting business going on. Their facial morphology is awesomely robust all with sagittal crests that served as attachments for the huge temporalis muscles, with what seems to be a pretty basic australopithecine both from the neck down. But here's the kicker, and something I honestly didn't know about three of the four robust guys we're dealing with here.
Their foramen magnum is heart shaped.

Yeah, my reaction too. Normally that feature is round, the point presents some interesting problems with the spinal cord and all of that but that would be pure speculation at this point.
To put it into perspective, here is a robust australopithecine next to a modern human skull.

And another with just the mandible.

To make it clear, the robusts are an evolutionary dead end, as in, they do not lead to modern humans. They did, however, live at the same time as early genus Homo.
Due to the stark contrast in the craniofacial morphology in the robust australopithecines, it's been proposed that they are different enough to deserve the genus name paranthropus. The jury is still out on that one for me.
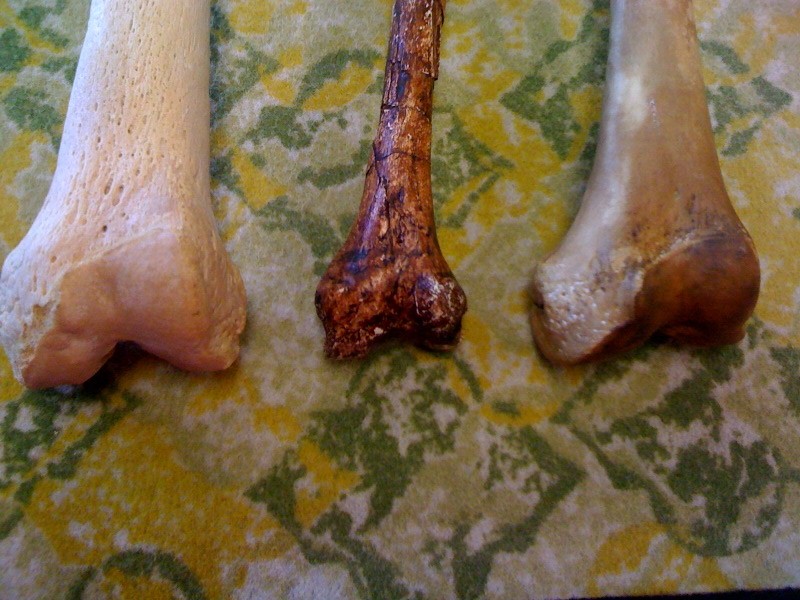
Due to the bias in the fossil record, we have a bunch of skull and mandibular bits from these guys but very little post-cranial pieces. We do have the pelvis from Australopithecus robustus, and it's quite small, definitely not of the magnitude of bone thickness that the cranium has. This is also from the only robust australopithecine without a heart shaped foramen magnum.

It seems that Australopithecus aethiopicus has a sort of incipient heart shaped foramen magnum, it isn't full blown but it definitely isn't round. My thought is that this gave rise to Australopithecus garhi and Australopithecus boisei, due to the similar morphology and the similar geography. The question is whether or not Australopithecus robustus has the extreme facial morphology due to similar environment or similar ancestry. Au. robustus differs in its geography (it hails from South Africa rather and East Africa) and it's lack of the heart shaped foramen magnum.

Here is the skull of Australopithecus boisei. You can clearly see VERY large zygomatics that flare out in order to accommodate a very large temporalis muscle. This muscle assists with bite force and must have been huge to need a sagittal crest to attach to in addition to the huge zygomatics. The environment was changing and the food was getting tougher so they needed to adapt. They accomplished this with increased bite force and tooth size.

The OH5 dental arcade, maxillary, showing how large the teeth are. Those molars are about four times the size of our own and are completely worn down to the dentin from the gnashing action accomplished by a huge masseter and temporalis. A consequence of adjusting the bite force was the loss of prognathism. It drew the face in so it was more flat, and therefore able to apply more force to the tough foodstuffs they were eating.